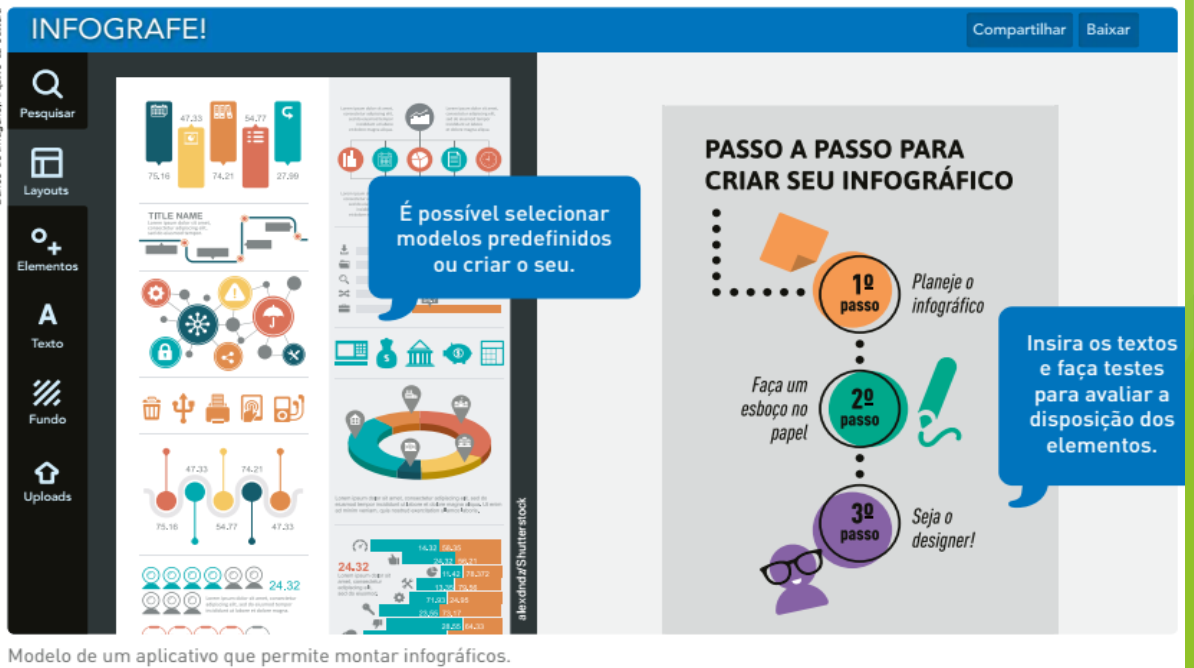
Uma das grandes contribuições da Ecologia para a compreensão da natureza foi a constatação de que as diversas espécies biológicas fazem parte de uma teia, na qual as comunidades de seres vivos se relacionam de diversas maneiras, que vão desde a competição até a cooperação entre espécies. O equilíbrio dos ecossistemas resulta exatamente dessa multiplicidade de relações ecológicas. Entre elas, uma das mais conhecidas e emblemáticas é a que se estabelece entre predador e presa. De tempos em tempos, certos animais são eleitos como astros de livros e filmes, despertando nas pessoas sentimentos de temor e admiração. Eles estão no topo da cadeia alimentar: são os superpredadores. Tigres, leões, leopardos, lobos e tubarões de olhos frios e assassinos são alguns dos predadores que escritores e cineastas mais utilizam para estimular a imaginação, despertando em nós sentimentos ancestrais, resquícios de uma época em que os antepassados dos seres humanos viviam em florestas e savanas povoadas por grandes carnívoros. Atualmente os grandes predadores estão de volta à cena, desta vez em artigos científicos que apontam o impacto ambiental causado pela crescente redução de suas populações. Um estudo internacional publicado recentemente pela revista científica Science indicou o declínio acentuado nas populações de grandes predadores. Ainda de acordo com o artigo, a perda de espécies no topo da cadeia alimentar devido à ação humana pode produzir impactos significativos nos ecos‑ sistemas terrestres.
porque aspectos importantes de seus nichos ecológicos – o tipo e o local de alimentação – são suficientemente distintos, o que evita uma competição acirrada.
2. Relações ecológicas intraespecíficas
Em uma comunidade biológica, os organismos mantêm relações tanto com seres de sua própria espécie quanto com os de outras espécies. Essas interações, genericamente denominadas relações ecológicas, podem ser classificadas em relações intraespecíficas, quando ocorrem entre indivíduos da mesma espécie, e relações interespecíficas, quando ocorrem entre indivíduos de espécies diferentes. Organismos de mesma espécie, por terem as mesmas necessidades fundamentais, quase sempre disputam os mesmos recursos do meio, fenômeno denominado competição intraespecífica. Há situações, entretanto, em que os membros da população se auxiliam mutuamente, trocando benefícios, o que se denomina cooperação intraespecífica. Competição intraespecífica Competição intraespecífica é a disputa entre indivíduos da mesma espécie por um ou mais recursos do ambiente. Dependendo da espécie, a competição pode ser por água, alimento, luz, locais para a construção de ninhos, parceiros para acasalamento etc. Além da luta física por alimento ou por parceiros de acasalamento, a competição intraespecífica também se manifesta de outras maneiras; por exemplo, se um indivíduo da espécie for muito ativo e eficiente na obtenção de alimento, ele tende a levar vantagem competitiva sobre outro menos ativo, sobretudo se o alimento for escasso. Plantas com sistemas radiculares desenvolvidos e eficientes, em determinado tipo de ambiente, podem levar vantagem sobre plantas menos dotadas na competição por água e por nutrientes (Fig. 3).
Figura 3 A competição por água entre plantas de desertos é evidenciada pela distribuição espaçada dos indivíduos da população. Na fotografia, deserto com plantas esparsas (Califórnia, EUA, 2019)
Cooperação intraespecífica
Em certas espécies, os indivíduos estabelecem relações cooperativas, o que se denomina cooperação intraespecífica. Esse tipo de cooperação desempenha um importante papel no sucesso evolutivo de muitas espécies. Entre as relações cooperativas intraespecíficas destacam ‑se o cuidado com a prole e a formação de grupos sociais. A cooperação intraespecífica manifesta ‑se claramente nas colônias e nas sociedades, que veremos a seguir.
Colônias
Do ponto de vista biológico, colônias são grupos de indivíduos de mesma espécie, fisicamente unidos, que interagem de forma mutuamente vantajosa, dividindo funções ou tarefas. As colônias podem variar em níveis de complexidade e no grau de divisão de tarefas entre os indivíduos. Algumas colônias são constituídas por indivíduos morfológica e funcionalmente semelhantes, sendo por isso denominadas colônias isomorfas (do grego isos, “igual”, “semelhante”, e morpho, “forma”). Um exemplo de colônia isomorfa é formada pela alga de água doce da espécie Volvox sp., que pode reunir mais de 50 mil indivíduos unicelulares e biflagelados dispostos lado a lado, formando uma esfera oca dentro da qual podem se formar colônias menores, que ficam protegidas por certo tempo no interior da “colônia ‑mãe” (Fig. 4)
Figura 4 (A) Fotomicrografia decolônias da alga verde Volvox aureus dentro de uma colônia ‑mãe.
(Microscópio fotônico; aumento q 63.) (B) Representação esquemática dos indivíduos biflagelados que formam a porção periférica das colônias. (Representação fora de proporção; cores meramente ilustrativas.)
Quando os indivíduos de uma colônia são diferentes morfológica e fisiologicamente, desempenhando funções distintas, fala‑se em colônia heteromorfa (do grego heteros, “diferente”). Um exemplo é o cnidário marinho Physalia physalis, popularmente conhecido como caravela ‑portuguesa. A caravela é formada por diversos tipos de indivíduos intimamente associados. Um dos indivíduos da colônia é uma bolsa cheia de gás, responsável pela flutuação; outros têm boca e cavidade digestiva, sendo responsáveis pela alimentação; outros são longos e ricos em células urticantes, formando tentáculos que atuam na captura de presas e na proteção da colônia; e há ainda indivíduos responsáveis pela reprodução. Na caravela ‑portuguesa, a especialização de funções faz com que os membros da colônia comportem ‑se quase como órgãos de um único organismo (Fig. 5).
Figura 5 (A) Fotografia do cnidário colonial Physalia physalis, constituído por indivíduos altamente integrados; a bolsa de gás pode atingir 30 cm de comprimento. (B) Representação esquemática que mostra os indivíduos que compõem uma caravela‑portuguesa. (Representação fora de proporção; cores meramente ilustrativas.) Sociedades
Sociedades são grupos de organismos de mesma espécie cujos indivíduos manifestam algum grau de cooperação, de comunicação e de divisão de trabalho, conservando, porém, relativa independência e mobilidade. É isso o que distingue sociedade de colônia: nesta última, como vimos no exemplo da caravela, os indivíduos são fisicamente unidos. Diversas espécies, entre as quais as formigas, as
abelhas e os seres humanos, vivem em sociedade (Fig. 6).
Figura 6 As abelhas melíferas (Apis mellifera) vivem em uma sociedade bem organizada, com divisão de funções entre os indivíduos. No centro da fotografia, pode ‑se observar a abelha ‑rainha, cercada por um séquito de abelhas operárias. A rainha mede cerca de 2,5 cm, enquanto as operárias medem 1,3 cm, em média.
3. Relações ecológicas interespecíficas
Os especialistas classificam as relações ecológicas entre espécies diferentes relações interespecíficas – em: competição interespecífica, interações tróficas, mutualismo, comensalismo e inquilinismo.
Competição interespecífica
Em seus experimentos, Gause observou que se duas espécies de uma comunidade disputam os mesmos recursos do ambiente, seus nichos ecológicos se “sobrepõem” parcialmente e ocorre competição interespecífica. Quanto mais os nichos ecológicos das espécies se assemelham em aspectos importantes, mais intensa é a competição; isso pode levar uma das espécies à extinção ou forçá‑la, a migrar em busca de uma área disponível e com menos espécies competidoras.
Interações tróficas
Herbivoria
Herbivoria é a relação que se estabelece entre as plantas e os animais que delas se alimentam. Certos animais comem apenas partes das plantas, não chegando a matá‑las. É o que ocorre, por exemplo, com mamíferos que se alimentam de grama e com larvas de insetos que comem folhas. Alguns animais se alimentam de sementes. Em qualquer dessas situações, do ponto de vista individual, há prejuízo
para as plantas e benefício para os animais que as consomem. A herbivoria é uma das mais importantes relações ecológicas da natureza, uma vez que, ao se alimentarem de plantas, os animais assimilam a energia captada da luz solar, que então é transferida para outros níveis tróficos das cadeias alimentares (Fig. 7)
Figura 7 A herbivoria é a relação pela qual a energia dos produtores, originalmente captada da luz, é aproveitada pelos consumidores primários. Na foto, capivara (Hydrochoerus hydrochaeris) com cerca de 120 cm alimentando ‑se de capim.
Predação
Predação, ou predatismo, é a relação em que uma espécie predadora mata e come indivíduos de outra espécie, sua presa. Do ponto de vista individual, a espécie predadora se beneficia, enquanto a espécie predada é prejudicada. Do ponto de vista ecológico, entretanto, a predação é um importante mecanismo regulador da densidade populacional, tanto de presas como de predadores. A estreita correlação entre as flutuações no tamanho das populações de predadores e presas é importante para a sobrevivência de ambas.
A importância da predação como mecanismo regulador das populações naturais ficou evidente no início do século XX, quando foi proibida a caça ao veado Odocoileus hemionus no Planalto de Kaibab, nos Estados Unidos. Ao mesmo tempo, estimulou‑se a perseguição aos predadores naturais dos veados – lobos, pumas e coiotes. Essas medidas fizeram com que a população de veados aumentasse rapidamente: em apenas 21 anos passou de 4 mil para 100 mil animais. Os campos de pastagem, porém, eram incapazes de suportar mais que 30 mil animais. Quando essa capacidade de suporte foi ultrapassada, os veados começaram a morrer de fome e de doenças e sua população diminuiu bruscamente. Quinze anos após ter atingido o recorde de 100 mil indivíduos, a população de veados reduziu ‑se a menos de 10 mil animais. Os veados, famintos, passaram a comer as plantas de capim até as raízes, além de pisotear excessivamente o solo, afetando a capacidade de recuperação das pastagens. Por isso o capim não voltou a brotar como antes, mesmo depois da redução drástica da população de veados (Fig. 8).
Figura 8 No gráfico, a linha azul mostra o crescimento da população do veado Odocoileus hemionus no Planalto de Kaibab, no Arizona, Estados Unidos, após proibição da caça e campanha de combate a seus predadores naturais. Se os predadores não tivessem sido eliminados, a expectativa seria de que o tamanho da população de veados seguisse a linha vermelha. À esquerda, foto do veado O. hemionus com um filhote (o animal adulto mede cerca de 1,5 m de comprimento).
Parasitismo
Parasitismo é uma relação ecológica em que uma espécie parasita vive à custa de uma espécie hospedeira, causando ‑lhe prejuízos. Em geral, as espécies parasitas e hospedeiras estão bem adaptadas umas às outras, de modo que a relação não causa prejuízos fatais ao parasitado. A tendência evolutiva de o parasita adaptar ‑se progressivamente ao hospedeiro, e vice‑versa, é denominada coadaptação. Organismos parasitas podem viver na superfície externa do hospedeiro, sendo chamados ectoparasitas (do grego ectos, “fora”), ou no interior do hospedeiro, sendo denominados endoparasitas (do grego endos, “dentro”). Exemplos de ectoparasitas são os piolhos e os carrapatos; exemplos de endoparasitas são as lombrigas, as solitárias, certas bactérias e os vírus (Fig. 9).
Figura 9 Exemplos de animais parasitas. (A) Carrapato da espécie Dermacentor variabilis, ectoparasita de animais, que mede aproximadamente 4 mm de comprimento. O parasita se alimenta do sangue do hospedeiro. (B) Solitária (Taenia sp.), verme endoparasita que habita o intestino humano e, que pode atingir alguns metros de comprimento. A tênia utiliza alimentos presentes no intestino do hospedeiro.
Parasitoidismo: parasitismo ou predação a longo prazo?
Um caso particular de parasitismo é o parasitoidismo, termo utilizado para designar a relação em que um organismo vive boa parte de sua vida dentro de um hospedeiro, parasitando ‑o. Em determinado momento, o hospedeiro é morto e consumido, como na predação. Exemplos bem conhecidos de parasitoides são as vespas que põem ovos dentro de hospedeiros como aranhas e larvas de insetos. Após a eclosão dos ovos, as larvas das vespas passam a devorar os tecidos corporais vivos do hospedeiro, até que ele seja consumido totalmente. Ao final do processo, as larvas transformam ‑se em uma fase denominada pupa, que origina a vespa adulta (Fig. 10).
Plantas parasitas
Há plantas que parasitam outras plantas. Um exemplo é o cipó‑chumbo, planta parasita de cor amarela, sem folhas nem clorofila, com aparência de fios de ovos, que cresce sobre outras plantas. Com suas raízes especializadas, o cipó ‑chumbo penetra no floema da planta hospedeira, de onde extrai seiva orgânica, da qual se alimenta. A erva ‑de ‑passarinho é outra planta que parasita outras plantas. Diferente‑ mente do cipó‑chumbo, a erva‑de‑passarinho tem folhas clorofiladas e produz moléculas orgânicas por fotossíntese, retirando da planta hospedeira apenas
água e sais minerais. Por extrair da hospedeira apenas substâncias inorgânicas, a erva ‑de ‑passarinho é considerada pelos botânicos hemiparasita (do grego hemi, “metade”) (Fig. 11).
Figura 10 Exemplo de parasitoidismo entre larvas de vespa da família Braconidae e lagarta de mariposa
da família Sphingidae. Após se desenvolverem no interior do corpo da lagarta, alimentando‑se de tecidos vivos, as larvas do parasita saem para a superfície do corpo do hospedeiro e tecem seus casulos, que podem ser vistos sobre a lagarta. Os casulos contêm pupas, que se transformam em vespas adultas.
Figura 11 Exemplos de parasitismo em plantas. (A) Erva ‑de ‑passarinho (Struthanthus flexicaulis), que pode ser identificada na fotografia por suas folhas maiores (medem cerca de 10 cm de comprimento) e mais escuras que as do jacarandá ‑mimoso (Jacaranda mimosaefolia, árvore com cerca de 15 m de altura), do qual extrai a seiva mineral. (B) O cipó ‑chumbo (Cuscuta sp.), de cor amarela e enrolado em uma planta hospedeira, extrai dela a seiva orgânica da qual se alimenta. A extração é feita por meio de raízes sugadoras denominadas haustórios.
Mutualismo
Mutualismo é uma relação ecológica em que ambas as espécies em interação obtêm benefícios. Os biólogos distinguem dois tipos de mutualismo: o facultativo e o obrigatório.
O mutualismo facultativo, também chamado de protocooperação, é uma relação ecológica em que as espécies associadas trocam benefícios, mas também podem viver sozinhas. A relação entre crustáceos do gênero Pagurus, os caranguejos‑eremitas, e algumas espécies de anêmona ‑do ‑mar (cnidários) é um exemplo de mutualismo facultativo. Esses animais não vivem necessariamente juntos, mas é frequente encontrá‑los em associação, o que é vantajoso para ambos. O caranguejo ‑eremita ocupa conchas vazias de caramujos, nas quais protege seu abdome delicado, pois, diferentemente de outros caranguejos, ele não tem carapaça rígida nessa região do corpo. Uma vez instalado em sua concha ‑moradia, o caranguejo ‑eremita a arrasta consigo para onde for, abandonando ‑a
apenas ao trocá‑la por outra maior. Sobre as conchas ocupadas por caranguejos‑eremitas é frequente haver fixadas uma ou mais anêmonas‑do‑mar; esses cnidários beneficiam ‑se da associação com o caranguejo por ganharem mobilidade e aproveitarem eventuais sobras de alimento do crustáceo. Este, por sua vez, se beneficia da capacidade defensiva das anêmonas ‑do ‑mar, cujos tentáculos têm células urticantes capazes de provocar queimaduras em eventuais inimigos. Outro exemplo de protocooperação é a relação entre alguns mamíferos – capivaras, búfalos, rinocerontes etc. – e aves que comem os carrapatos que os infestam. Há vantagens tanto para o mamífero, que se livra dos incômodos ectoparasitas, quanto para o pássaro, que obtém alimento com relativa facilidade.
Crocodilos também convivem cooperativamente com aves que entram em sua boca, de onde removem detritos e sanguessugas presos às gengivas (Fig. 12).
Figura 12 Exemplos de protocooperação. (A) Representação esquemática de um caranguejo‑eremita colocando deliberadamente uma anêmona‑do‑mar sobre a concha onde mora. (Representação fora de proporção; cores meramente ilustrativas.) (B) Caranguejo‑eremita (Dardanus calidus) dentro da concha que lhe serve de abrigo, sobre a qual há anêmonas ‑do ‑mar da espécie Calliactis parasitica. O caranguejo mede cerca de 12 cm. (C) Capivara (Hydrochoerus hydrochaeris), que chega a mais de 1,30 m de comprimento, tendo sobre a cabeça a ave suiriri ‑cavaleiro (Machetornis roxisus, com cerca de 18 cm de comprimento), que se alimenta de carrapatos presentes na capivara. (D) Pássaro (Pluvianus aegyptius, em média 20 cm de comprimento) que se alimenta de detritos e eventuais parasitas presentes na boca do crocodilo (Crocodylus niloticus, que atinge cerca de 4 m de comprimento).
Mutualismo obrigatório é um tipo de relação permanente e indispensável à sobrevivência dos indivíduos associados. Um exemplo desse tipo de mutualismo é a interação entre certas espécies de cupim e microrganismos – bactérias e protozoários – que habitam seu intestino. Os cupins alimentam ‑se de madeira, mas são incapazes de digerir seu principal componente, a celulose. A digestão desse carboidrato é realizada por microrganismos que vivem exclusivamente no tubo digestivo desses insetos. Outro exemplo de mutualismo obrigatório são os liquens, formados pela associação de certas espécies de algas verdes ou de bactérias fotossintetizantes com determinadas espécies de fungos. Por meio da fotossíntese, a alga ou a bactéria produz matéria orgânica, utilizada pelo fungo; este, por sua vez, é capaz de absorver do ambiente água e nutrientes, que seus parceiros fotossintetizantes aproveitam. O líquen sobrevive em locais onde nenhuma das duas espécies que o formam poderia sobreviver isoladamente.
Um terceiro exemplo de mutualismo obrigatório é a micorriza (do grego, mycos, “fungo”, e rhizos, “raiz”),associação entre determinados fungos e raízes de plantas. As hifas dos fungos penetram nas raízes, das quais obtêm carboidratos como glicose e sacarose. A planta, por sua vez, beneficia ‑se da grande capacidade do fungo de absorver água e nutrientes minerais do solo (Fig. 13).
Figura 13 Exemplos de mutualismo. (A) Certas espécies de cupim, como a da foto (Zootermopsis angusticollis, com cerca de 1 mm de comprimento), abrigam em seu tubo digestivo bactérias e protozoários flagelados. Ao lado da foto, representação esquemática do protozoário do gênero Trichonympha, que vive no intestino de cupins. (B) Liquens das espécies Xanthoria parietina e Physcia tenella sobre o tronco de uma árvore. Esses liquens medem cerca de 10 cm de diâmetro. Ao lado da foto, representação esquemática da relação entre células de uma alga verde e o fungo no líquen. (C) Fotomicrografia de raízes de soja (Glycine max) envoltas por fungos formando micorrizas. (Microscópio eletrônico de varredura; aumento q 150×; cores meramente ilustrativas.) Ao lado da foto, representação esquemática da associação de um fungo com as raízes de uma planta na micorriza. A planta recebe água e sais minerais que o fungo extrai do solo; o fungo utiliza substâncias orgânicas produzidas pela planta. (Ilustrações à direita fora de proporção; cores meramente ilustrativas.)
Comensalismo e inquilinismo
Comensalismo é o tipo de relação em que uma das espécies associadas é beneficiada, enquanto a outra não tem prejuízos nem benefícios. O principal recurso buscado pelo comensal, como o nome
sugere, é alimento. Um exemplo conhecido de comensalismo é a associação entre a rêmora (ou peixe‑piloto) e o tubarão. A rêmora tem uma estrutura dorsal aderente, semelhante a uma ventosa, com a
qual se prende ao corpo do tubarão. Este proporciona transporte e restos de alimento para a rêmora e parece não se importar com sua presença. A relação entre abutres e animais carnívoros também é um tipo de comensalismo. Os abutres acompanham os carnívoros a distância, servindo ‑se dos restos da caça deixados pelos predadores (Fig. 14). Inquilinismo é a relação em que uma espécie “inquilina” vive sobre uma espécie hospedeira, ou dentro dela, sem prejudicá‑la. O principal recurso buscado pelo inquilino, como o nome sugere, é abrigo e moradia. Várias espécies de orquídeas, bromélias e samambaias vivem sobre outras plantas maiores que lhes servem de
suporte. Nesses casos, as espécies inquilinas são denominadas epífitas (do grego epi, “sobre”, e phytos, “planta”). A vantagem de crescer sobre plantas mais altas é obter maior suprimento de luz para a fotossíntese, principalmente em ambientes pouco iluminados, como o interior das florestas.
Figura 14 Exemplos de comensalismo. (A) Rêmoras (peixes da família Echeneidae) “pegam carona” em um tubarão‑limão (Negaprion brevirostris). O tubarão‑limão pode chegar a 3 m de comprimento e as rêmoras, a 90 cm. (B) Abutres‑de‑costas‑brancas (Gyps africanus) são comensais habituais de felinos como os guepardos (Acinonyx jubatus) e outros carnívoros africanos. Os abutres e os guepardos medem, respectivamente, cerca de 80 cm e 1,3 m de comprimento.
O conceito de simbiose
Em 1879 o biólogo alemão Heinrich Anton de Bary (1831 ‑1888) criou o termo simbiose (do grego syn, “juntos”, e bios, “vida”) para designar associações entre indivíduos de espécies diferentes com consequências vantajosas ou desvantajosas para pelo menos uma das partes. Segundo a definição original, relações como inquilinismo, comensalismo, mutualismo e parasitismo seriam casos de simbiose. No entanto, o termo simbiose tem sido utilizado frequentemente como sinônimo de mutualismo. Além disso, alguns autores definem simbiose como uma associação entre indivíduos de espécies diferentes em que um vive dentro do outro ou sobre o outro. Por isso, é preciso estar atento à utilização do termo.
Resumo das relações ecológicas interespecíficas
Apresentamos a seguir um resumo (Tab. 1) das relações interespecíficas estudadas neste capítulo. Os efeitos da relação entre os indivíduos das espécies relacionadas são representados pelo sinal (+), quando há ganho ou vantagem, pelo sinal (-), quando há perda ou desvantagem, e pelo número (0), quando não há vantagem nem desvantagem. Lembre‑se de que esse critério visa apenas caracterizar a relação em termos de perdas ou ganhos individuais. Como enfatizamos no capítulo, relações como a predação, a competição e o parasitismo, em que há prejuízos para os indivíduos, podem trazer benefícios para populações, comunidades e ecossistemas.
Nem sempre é fácil caracterizar as relações entre os seres vivos. Os biólogos procuram utilizar determinados critérios lógicos para facilitar esse estudo, mas nem sempre eles correspondem à diversi‑ dade da natureza. Em certos casos, é difícil estabelecer a diferença entre inquilinismo e comensalismo. Certas espécies de peixe ‑palhaço, por exemplo, encontram abrigo e proteção entre os tentáculos de anêmonas‑do‑mar. Essa relação seria caracterizada, em princípio, como inquilinismo. Entretanto, se os peixes‑palhaço se aproveitam de restos da alimentação da anêmona, além de utilizá‑la como abrigo, poderíamos classificar a relação como comensalismo? Pense nisso.
As populações naturais interagem dinamicamente em suas comunidades biológicas. A trama ecológica, construída ao longo de milhões de anos de evolução, recentemente tem sofrido fortes influências antrópicas ou, em termos menos técnicos, da espécie humana; anthropus, em grego, significa "ser humano". O grande aumento da população humana mundial agrava cada vez mais os desequilíbrios ecológicos. Biomas inteiros estão sob ameaça de extinção. Somente a compreensão da natureza e o desenvolvimento de uma consciência ecológica, com foco na sustentabilidade, poderão nos ajudar a resolver as graves crises ambientais que se avizinham. Os dados sobre o crescimento da população humana são realmente impressionantes. Segundo estimativas recentes, a população mundial atingiu 7,7 bilhões de pessoas em abril de 2019. Em 1950 eram “apenas” 2,5 bilhões. Ou seja, o número de seres humanos no planeta
mais que triplicou nos últimos 69 anos e poderá ultrapassar os 11,2 bilhões em 2100. Segundo dados do Fundo de Populações das Nações Unidas, o primeiro bilhão de pessoas foi atingido em 1804. A população levou 123 anos para dobrar de tamanho e atingir os 2 bilhões, o que ocorreu em 1927. Cerca de 47 anos depois, em 1974, a população atingiu a cifra de 4 bilhões de pessoas. Em 2019, já éramos nada menos que, 7,7 bilhões. Os demógrafos estimam que a humanidade contará com 8 bilhões de pessoas por volta de 2024. Em 1798 o demógrafo e economista britânico Thomas Malthus (1766-1834) teorizou que haveria fome e crise social no futuro se o crescimento populacional não fosse freado. As previsões de Malthus não consideraram, porém, que haveria progressos científicos e tecnológicos capazes de aumentar consideravelmente a produção de alimentos, além de outros fatores, como a evolução dos métodos de controle da natalidade, a diminuição do número de filhos por mulher
e o planejamento familiar. A principal crítica a Malthus e a seus seguidores mais modernos, os neomalthusianos, é a visão simplista de que a pobreza e a fome no planeta decorrem apenas do tamanho da população, e não da má distribuição das riquezas. Além disso, há uma corrente de pensamento que defende exatamente o contrário: a superpopulação é que é consequência direta da pobreza, da falta de informação e de melhores condições de vida dos povos do mundo. Estudar as populações biológicas é importante porque elas são unidades funcionais básicas nos ecossistemas, estando na base dos sistemas ecológicos. O estudo de certas características de uma população ao longo do tempo informa se ela está em expansão, em declínio ou estável, o que permite estabelecer correlações com fatores como disponibilidade de alimento e clima, entre outros. Ainda há muitas divergências acerca do crescimento da população humana e sua relação com a pobreza e a desigualdade social no mundo. Como já foi dito, alguns estudiosos consideram que a miséria e a fome no mundo estão mais relacionadas às más políticas de distribuição de recursos do que à superpopulação. Entretanto, não há dúvida de que muitos problemas da humanidade se agravam quando o ambiente natural é degradado, o que é consequência direta da quantidade de pessoas que o exploram. Neste capítulo, estudaremos, além das populações, a sucessão ecológica, processo pelo qual populações e comunidades se sucedem e evoluem durante a colonização de uma área. Diferentes tipos de comunidades atingem seu clímax em diversos locais do planeta: cerrados, savanas, florestas, desertos etc. A valorização e a preservação desses ambientes naturais são grandes desafios atuais e futuros para a humanidade. Reconhecer as feições características de cada ambiente e estar informado sobre o crescimento da população humana são ações pela cidadania.
1. Características das populações biológicas
O conceito de população
População biológica é definida como um conjunto de indivíduos de mesma espécie que convivem em determinada área e que podem cruzar entre si de forma natural. Vamos partir de um exemplo hipotético em que duas espécies distintas, uma de pássaros e uma de ratos, vivem em diversas ilhas de um arquipélago. Os pássaros, capazes de voar livremente entre as ilhas, podem se encontrar e cruzar entre si. De acordo com os biólogos, esses pássaros formam uma única população, distribuída pelas ilhas. Suponhamos que os ratos não consigam nadar entre as ilhas; consequentemente, ratos das diferentes ilhas do arquipélago não se cruzam, e cada ilha teria uma população particular desses roedores. Aplicando o mesmo critério à espécie humana, pode-se dizer que atualmente a humanidade constitui uma única população biológica, que se distribui pelos diversos continentes. As migrações, facilitadas principalmente pela revolução nos transportes e nas comunicações, transformaram a população humana em uma única “aldeia global”, termo criado pelo filósofo canadense Herbert Marshall McLuhan (1911-1980) nos anos 1960. Na linguagem cotidiana, por simplificação, é comum referir-se à população de um município, de um estado, de um país etc., quando o mais correto seria utilizar o termo subpopulação. O globalismo da
população humana ficou tristemente evidenciado com o rápido alastramento da pandemia da COVID-19, originada em uma pequena cidade na China e que em poucos meses se espalhou por quase 200 países, em todos os continentes.
Densidade populacional
Uma informação importante a respeito de uma população é a densidade populacional, definida como a relação entre o número de indivíduos da espécie e a área ou o volume (no caso de hábitats aquáticos, por exemplo) que eles ocupam. Essa definição está formulada a seguir: Densidade populacional 5 No de indivíduos 4 Área ou volume Na população humana, o estudo estatístico do tamanho populacional e de sua composição em idade e sexo, entre outros aspectos, constitui a demografia (do grego demos, “povo”, e graphe, “descrição”). Quando se trata de populações humanas, fala-se em densidade demográfica, calculada com base em levantamentos periódicos conhecidos como censos demográficos. O censo demográfico realizado no Brasil em 1990, por exemplo, estimou a população brasileira em aproximadamente 150 milhões de pessoas, distribuídas pelos 8,5 milhões de quilômetros quadrados de superfície do território nacional. Assim, a densidade demográfica do Brasil em 1990 era de aproximadamente 17,6 hab/km2 (habitantes por quilômetro quadrado). Em 2000, o censo mostrou que a população brasileira tinha aumentado para 169 milhões de pessoas; como o território permaneceu o mesmo, a densidade demográfica brasileira cresceu para 19,8 hab/km2. Em 2006 o Instituto Brasileiro de Geografia e Estatística (IBGE)* estimou a população brasileira em 186 milhões de habitantes e, em 2010, em 190.732.694 pessoas, o que indica que nesta última data a densidade demográfica era de aproximadamente 22,4 hab/km2. Em 2019 a população brasileira foi estimada em 208,5 milhões de pessoas, o que eleva a densidade demográfica para 24,5 hab/km2, um aumento de quase 40% em relação a 1990 (Fig. 1).
Figura 1 No Brasil há desde centros urbanos superpovoados até áreas rurais pouco ocupadas. (A) Rua 25 de Março, na cidade de São Paulo, SP, 2013. (B) Fazenda na zona rural de Tiradentes, MG, 2019.
Taxas populacionais
Um aspecto importante em Ecologia é saber como uma população cresce. Para isso, estima-se a taxa de crescimento populacional, que é a variação (aumento ou diminuição) do número de indivíduos em determinado intervalo de tempo. Vamos concretizar essa definição analisando os dados da Tabela 1, na coluna lateral (abaixo), relativos a populações hipotéticas de bactérias. Para determinar a taxa de crescimento das populações, primeiramente calculamos a variação (aumento ou diminuição) do número de indivíduos em um intervalo de tempo considerado. Para isso, tomamos o número de indivíduos da população no tempo final (Nf) e subtraímos dele o número de indivíduos no tempo inicial (Ni). Essa variação bruta é então dividida pelo número de indivíduos que havia na população no tempo inicial (Ni). Finalmente esse número é dividido pela duração do período considerado (t). Veja a fórmula empregada no cálculo:
Você pode estar se perguntando se esses cálculos são necessários. Note que, ao dividirmos a variação bruta pelo tamanho inicial da população, estamos relativizando-a, isto é, levando em conta não os números brutos, mas o aumento do número de indivíduos em relação ao tamanho inicial da população. No exemplo, a população A teve um acréscimo de apenas 30 mil indivíduos, enquanto a população B teve um acréscimo de 300 mil indivíduos no tempo considerado. Isso indica que a população B cresceu mais que a A? Levando em conta o tamanho inicial da população, concluímos que não; ao contrário, a população A teve uma taxa de crescimento maior do que a da população B, se considerarmos que o tamanho inicial da população A era menor do que o da população B. Por levar em conta o tamanho inicial da população, fala-se em taxa de crescimento relativo (TCR) da população. Aplicando a fórmula, as taxas de crescimento relativo (TCR) para as duas populações de bactérias são:
Portanto, a taxa de crescimento da população A, no período analisado, foi o dobro da taxa de crescimento da população B.
Taxas de natalidade e de mortalidade
O crescimento de uma população é determinado, fundamentalmente, por dois fenômenos de efeitos opostos: a natalidade, ou seja, o número de indivíduos que nascem, e a mortalidade, o número de indivíduos que morrem. Outros fatores que também afetam o tamanho de uma população são a imigração – a entrada de novos indivíduos na população, e a emigração, a saída de indivíduos da população. Quando se trata da população humana, a taxa de natalidade é definida como o número de crianças nascidas no período de um ano para cada 1.000 habitantes da população. Analogamente, a taxa de mortalidade é o número de óbitos (mortes) ocorridos no período de um ano para cada 1.000 habitantes da população. Por que expressar o número de nascimentos para cada 1.000 habitantes, isto é, dividir o número de nascimentos por 1.000? O motivo é o mesmo que utilizamos ao dividirmos um número por 100 quando expressamos porcentagens. O que estamos fazendo, nos dois casos, é relativizar os números brutos, o que facilita compará-los aos de outras populações. Por exemplo, se ocorrerem dois nascimentos por ano em uma população de 1 mil habitantes em determinado ano, a taxa de natalidade é igual a 2 ÷ 1.000, ou 0,002. Em outra população, de 4 mil habitantes, nasceram quatro pessoas nesse mesmo ano; nesse caso, a taxa de natalidade foi de 4 ÷ 4.000, ou 0,001, a metade da taxa da primeira população. As taxas de mortalidade também podem ser detalhadas por faixa de idade. Quando se fala em taxa de mortalidade infantil, por exemplo, divide-se o número de óbitos de crianças com menos de 1 ano por 1.000 crianças nascidas vivas no período de 1 ano.
Figura 2 Curva de crescimento intrínseco de uma população de bactérias que duplica de tamanho a cada hora e tem índice de mortalidade zero no período considerado. Gráficos com curva semelhante são esperados para qualquer população biológica. Esse tipo de curva é característico de um crescimento em progressão geométrica, em que, a intervalos iguais de tempo, o número de indivíduos da população dobra.
Índice de fertilidade
Uma informação importante sobre certos tipos de população é seu índice de fertilidade, ou taxa de fertilidade, definido como o número médio de descendentes que uma fêmea gera durante seu período reprodutivo. No caso da espécie humana, se um país tiver índice de fertilidade igual a 2, significa que o número médio de filhos por mulher é 2. Espera-se, então, que os filhos de cada geração substituam seus pais, o que tende a manter o tamanho da população estável. Se o índice de fertilidade for superior a 2, há uma tendência
ao crescimento populacional; se for inferior a 2, há uma tendência à diminuição do tamanho da população, situação denominada crescimento negativo.
Crescimento populacional
A maioria das populações tem grande potencial para crescer. Se a mortalidade fosse zero, uma única bactéria, reproduzindo-se a cada 20 minutos, produziria descendência suficiente para cobrir a Terra em apenas 36 horas. Um único paramécio poderia gerar, em alguns dias, uma massa de indivíduos correspondente a 10 mil vezes a massa da Terra. Um único casal de pássaros e seus descendentes, chocando de 5 a 6 ovos por ano, produziria 10 milhões de descendentes em 15 anos. Essa capacidade teórica de crescimento de uma população biológica é denominada curva de crescimento intrínseco, ou potencial biótico. Em populações teóricas, se representarmos em um gráfico o número de indivíduos da população em função do tempo, a curva resultante é exponencial e corresponde ao crescimento intrínseco, ou também chamado de potencial biótico da população (Fig. 2).
Em condições naturais, o potencial de crescimento de uma população é limitado pela disponibilidade de recursos do meio (alimentos, espaço, abrigo etc.) e pela ação de predadores, parasitas e populações competidoras. A ação desses fatores em conjunto constituem a resistência do meio, geralmente levando as taxas de natalidade e de mortalidade a se equivalerem, de modo que o número de indivíduos de uma população tende a permanecer mais ou menos constante ao longo do tempo. Para uma população em particular, há um limite máximo de indivíduos que o ambiente consegue suportar, denominado capacidade de suporte, ou carga biótica máxima do meio. O crescimento de uma população a partir de alguns poucos indivíduos, se for representado em um gráfico que relaciona o número de indivíduos em função do tempo, resulta em uma curva ascendente em forma da letra S, que tende a se estabilizar quando é atingida a capacidade de suporte do meio. O aspecto apresentado
pela curva de crescimento populacional resulta da interação entre o crescimento intrínseco e a resistência do meio, esta última, como vimos, constituída pelo conjunto de fatores ambientais que limitam o crescimento da população (Fig. 3).
Figura 3 Gráfico que representa a curva de crescimento de uma população originada de um pequeno número de indivíduos.
2. Fatores que regulam o tamanho populacional
O crescimento de uma população biológica é limitado pela densidade populacional, pela disponibilidade de alimento, pela competição com indivíduos de outras espécies (competição interespecífica), pela predação e pelo parasitismo, entre outros fatores. O estudo de certas populações mostrou que quando a densidade populacional aumenta além de certo limite, a taxa de natalidade tende a diminuir. Essa situação ocorre, em geral, como consequência do aumento da competição por alimento e da competição entre os casais por locais de procriação. Em experimentos com ratos, verificou-se que quando as gaiolas de criação se tornavam superpovoadas, mesmo com alimento em abundância, a taxa de natalidade caía a zero. Muitos filhotes chegavam mesmo a morrer antes do nascimento, ainda no útero materno, o que levou à conclusão de que essas mortes estariam relacionadas ao estresse e à tensão emocional decorrentes do superpovoamento. Quanto mais intensos são os fatores que limitam o crescimento, menor é o aumento populacional. Em contrapartida, o abrandamento de um ou mais fatores limitantes pode levar ao aumento da taxa de crescimento da população. Como a intensidade dos fatores limitantes geralmente varia ao longo do tempo, observa-se que o tamanho das populações passa por flutuações periódicas (Fig. 4). Um exemplo clássico da regulação do tamanho populacional por meio da predação é o da flutuação dos tamanhos de populações de linces e lebres da região ártica do Canadá. Os dados foram coletados durante 80 anos (de 1855 a 1935) pela Companhia da Baía de Hudson, que registrava o número de peles comercializadas pelos caçadores da região. Como o número de caçadores era conhecido e
sofria pouca alteração de um ano para o outro, as variações na quantidade de peles comercializadas provavelmente refletiam as variações do tamanho relativo das populações das espécies caçadas.
Traçando no mesmo gráfico as curvas de densidade populacional de lebres e de linces, verifica-se que a população de linces geralmente alcança seu maior desenvolvimento um a dois anos depois que a população de lebres atinge seu ápice. Uma interpretação bastante plausível é que o tamanho das populações de lebres e de linces depende da relação presa-predador existente entre essas duas espécies. Quando a população de lebres aumenta, a população de linces também aumenta, já que há maior oferta de alimento para o predador. Com o aumento do número de linces, intensifica-se a predação, o que leva à diminuição da população de lebres. Com menos lebres para caçar, a população de linces volta a diminuir, o que permite a retomada do crescimento da população de lebres, e assim por diante.
Figura 4 Gráfico que mostra a curva de crescimento da população de carneiros na Austrália, estimada com base em dados coletados entre as décadas de 1830 e 1930. A linha azul representa a população real, cujo tamanho oscila em decorrência das variações de fatores ambientais que regulam o crescimento populacional. A linha vermelha, denominada curva logística, representa o crescimento teórico de uma população que se estabiliza ao atingir a capacidade de suporte do meio (seta).
Figura 5 (A) Gráfico que mostra as flutuações no tamanho das populações de lebres (em azul) e de linces (em vermelho) no Canadá, entre 1855 e 1935. (B) Foto de lince (Lynx sp.) predando uma lebre branca (Lepus sp.). O lince e a lebre medem, respectivamente, cerca de 1 m e 50 cm de comprimento.
Fonte: adaptado de DAJOZ, R. Ecologia geral. 3. ed. São Paulo: Edusp, 1978.
FOTO: VINICIUS BACARIN/SHUTTERSTOCK; ILUSTRAÇÃO: ERICSO
A explosão populacional da humanidade
A população humana é semelhante a outras populações biológicas e está sujeita aos mesmos fatores gerais que regulam e limitam o crescimento populacional de outras espécies. Entretanto, a humanidade tem conseguido controlar alguns fatores ambientais, o que permitiu um formidável ritmo de crescimento populacional. O crescimento acelerado das populações humanas deve-se principalmente à diminuição da taxa de mortalidade, decorrente tanto dos avanços agrícolas e tecnológicos, que contribuíram para o aumento da produção de alimentos, como dos progressos médicos e sanitários, que prolongaram a expectativa de vida (Fig. 6)
Figura 6 O crescimento acelerado da população humana nas últimas décadas tem levado os cientistas a fazer diversas projeções sobre suas possíveis curvas de crescimento (A ou B, mostradas no gráfico). Na fotografia ao fundo, a Avenida Paulista, na capital de São Paulo, em um domingo sem tráfego de automóveis, em 2019.
Um problema inerente à expansão demográfica ilimitada é a necessidade de produzir cada vez mais alimentos para nutrir o número crescente de pessoas. Não é possível ampliar indefinidamente as áreas de terra cultivada. A maioria das terras férteis já está sendo utilizada, e muitas delas já tiveram seus recursos
esgotados; certas áreas demandariam tanto empenho para se tornar produtivas que, pelo menos por enquanto, não é vantajoso explorá-las. Além disso, os ecossistemas naturais que ainda restam, por exemplo, os da Amazônia, dos Cerrados e do Pantanal Mato-Grossense, não devem ser explorados de forma predatória. É preciso manter áreas preservadas para que a diversidade biológica, ou biodiversidade, produzida ao longo de bilhões de anos de evolução não seja perdida. Biomas como a Floresta Amazônica e o Cerrado têm forte influência na dinâmica de chuvas e rios que abastecem grande parte do continente, e sua devastação teria consequências catastróficas para a biosfera. Até a década de 1970, certos países consideravam o controle da natalidade uma iniciativa racista, reacionária ou imperialista. Hoje, os cidadãos da maioria das nações consideram importante realizar algum tipo de controle da natalidade e o planejamento familiar para manter a qualidade de vida da população. A maioria dos países desenvolvidos já conseguiu frear o crescimento populacional, o que também tem ocorrido em alguns países em desenvolvimento, como a Tailândia, a Colômbia e a Costa Rica, que já alcançaram reduções significativas de suas taxas de natalidade. Outros, apesar das tentativas, ainda não alcançaram o objetivo desejado. Embora os problemas da humanidade sejam decorrentes de vários fatores, imagina-se que se o crescimento da população for freado, será possível ganhar tempo para resolver problemas que seriam agravados em um quadro de superpopulação, como a fome, as desigualdades econômicas, a degradação ambiental e várias doenças. O escritor estadunidense Lester R. Brown (1934-) iniciou um de seus livros, O vigésimo nono dia, publicado em 1980 (Editora FGV), com uma pequena história: “Para ensinar às crianças a noção de crescimento exponencial, os professores franceses se valem de uma charada. Em uma lagoa flutua uma folha de árvore. A cada dia que passa, o número de folhas dobra: duas folhas no segundo dia, quatro no terceiro, oito no quarto, e assim por diante. Se a lagoa ficar inteiramente coberta de folhas no trigésimo dia, quando ela ficou coberta pela metade? Resposta: No vigésimo nono dia”. A lagoa corresponde ao nosso planeta e as folhas, às pessoas que o habitam. Talvez o planeta já esteja coberto pela metade e logo se torne repleto de gente. O maior risco dessa situação é ignorar os sinais da iminente saturação ou interpretá-los de modo errôneo. Como o trigésimo dia poderá não oferecer possibilidades de sobrevivência, é importante não chegar a ele; alguns propõem que um dos caminhos para isso seja limitar o crescimento demográfico.
Paradoxalmente, o aumento demográfico também está ligado ao grau de desenvolvimento da saúde pública. A população cresce não apenas porque há mais nascimentos, mas também porque a duração média da vida humana tem aumentado. A expectativa de vida em 1650 era de 30 anos; hoje, a média mundial é de mais de 65 anos e, em países desenvolvidos, ultrapassa os 80 anos. De acordo com dados coletados em 2018 pelo IBGE, a expectativa de vida média no Brasil já passa dos 76 anos. Veja na figura a seguir uma comparação entre as pirâmides de idade da Nigéria, do Brasil e da Itália (Fig. 7).
Figura 7 Pirâmides de idade da Nigéria, do Brasil e da Itália para dados de 2020. A base larga e o ápice estreito da pirâmide da Nigéria indicam, respectivamente, alta taxa de natalidade e, provavelmente, alta taxa de mortalidade nas idades mais avançadas, o que dá um aspecto bem triangular ao gráfico. Na pirâmide da Itália, típica de países desenvolvidos, o gráfico tem base mais estreita e ápice mais largo, indicando, respectivamente, controle da natalidade e expectativa de vida elevada. O que se pode dizer sobre a pirâmide do Brasil?
Fonte: adaptado de ORGANIZAÇÃO DAS NAÇÕES UNIDAS. Disponível em: <https://population.un.org/wpp/>.
Acesso em: 28 abr. 2020
3. Sucessão ecológica
Os biólogos denominam sucessão ecológica o processo ordenado de alteração nas comunidades durante a colonização de um ambiente. O ambiente colonizado pode ser inicialmente desprovido de vida ou já ter sido anteriormente ocupado por uma comunidade biológica, destruída por algum desastre natural ou pela atividade humana (antrópica). No decorrer do processo de sucessão, as espécies que compõem a comunidade biológica alteram-se gradualmente de forma ordenada e progressiva. Quando a sucessão ocorre em ambientes inicialmente desprovidos de vida, onde as condições de clima e de solo são pouco favoráveis ao estabelecimento de seres vivos, como superfícies formadas por lavas vulcânicas recém-solidificadas, rochas e dunas, fala-se em sucessão ecológica primária. Apesar das condições iniciais altamente desfavoráveis, certas espécies de organismos – as espécies pioneiras – conseguem se instalar nesses lugares inóspitos e abrir caminho para a chegada de outras espécies.
Dunas de areia, por exemplo, costumam ser colonizadas por certas espécies de gramíneas, cujas sementes são transportadas pelo vento. Essas gramíneas pioneiras conseguem suportar o calor, a escassez de água e o solo pouco estável e iniciam a colonização da duna. Ao longo das gerações de gramíneas, cada vez mais matéria orgânica se acumula no solo, favorecendo a sucessão ecológica de novas comunidades. Superfícies de rochas nuas são geralmente colonizadas por liquens, que se fixam sobre as rochas e originam uma camada orgânica, que pode abrigar mais tarde musgos e gramíneas, dando início à sucessão ecológica. Os biólogos utilizam o termo ecese para designar a comunidade inicial de uma sucessão ecológica (Fig. 8).
Quando a sucessão ocorre em locais parcialmente destruídos, mas anteriormente ocupados por comunidades biológicas, fala-se em sucessão ecológica secundária. Exemplos de sucessão ecológica secundária são a recolonização de campos de cultivo abandonados, de regiões desflorestadas, de áreas destruídas por queimadas ou de
lagos recém-formados. Nesses casos as mudanças geralmente são mais rápidas do que na sucessão primária, uma vez que as condições iniciais já são mais favoráveis ao estabelecimento de seres vivos. À medida que o ambiente é colonizado pelas espécies pioneiras, ele vai gradualmente se modificando. Ocorrem variações menos bruscas na temperatura do solo, e o grau de umidade tende a aumentar. O material orgânico resultante da decomposição dos cadáveres das espécies pioneiras acumula-se no solo, disponibilizando nutrientes e favorecendo a retenção de água. No caso das dunas, por exemplo, as raízes das plantas pioneiras contribuem para estabilizar o solo, evitando que as partículas arenosas sejam facilmente carregadas pelo vento. As novas condições criadas pelas espécies pioneiras favorecem a chegada de outras plantas e de animais, que podem se estabelecer no local. As espécies recém-chegadas tendem a competir com as pioneiras e pouco a pouco tomam o lugar delas. As sucessivas gerações de plantas e animais que nascem, crescem, morrem e se decompõem em um local de sucessão tornam o solo cada vez mais rico em matéria orgânica e umidade, e em uma área antes desabitada passa a haver uma comunidade biológica cada vez mais estável. A complexidade da comunidade depende de vários fatores, entre eles o tempo que se passou desde o início da colonização, as condições climáticas do ambiente no local da sucessão, as espécies colonizadoras e as que se estabeleceram no processo (Fig. 9). A cada estágio do processo de sucessão, a comunidade contribui para alterar cada vez mais a estrutura do ambiente e as condições climáticas locais, surgindo assim um microclima, ou seja, um clima próprio da comunidade em sucessão,
que traz novas possibilidades para a colonização por outras espécies. Ao longo da sucessão, as comunidades que se formam têm aumento na biodiversidade, na biomassa e na estabilidade das relações ecológicas. O processo de sucessão ecológica leva ao estabelecimento de uma comunidade biológica relativamente estável e bem adaptada às condições locais. A estabilidade depende da rede de interações construídas ao longo da sucessão das comunidades, com o passar do tempo. A comunidade que surge no estágio final de sucessão é denominada comunidade clímax, nela, a biodiversidade, a biomassa e as condições microclimáticas tendem a se estabilizar ao longo do tempo. As comunidades intermediárias em um processo de sucessão ecológica são chamadas de seres ou estágios serais da sucessão.
.
Figura 8 (A) Dunas no Parque Estadual do Jalapão, Mateiros, TO (2019). (B) Liquens sobre rochas na Chapada do Araripe, Santana do Cariri, CE (2020)
Figura 9 Representação esquemática de sucessão ecológica em dunas de areia. Nesse caso, a vegetação pioneira contribui para a fixação das dunas, o que modifica a atuação dos ventos e traz novas alterações climáticas. O clima torna-se cada vez mais propício ao estabelecimento de seres vivos. (Representação fora de proporção; cores meramente ilustrativas.)
4. Os grandes biomas do mundo
O conceito de bioma
Pode-se definir bioma (do grego bio, “vida”, e oma, “grupo”) como uma área geográfica na qual se desenvolve um ecossistema relativamente uniforme, caracterizado pelo clima, pelo solo e pela fisionomia de sua vegetação. Regiões da Terra com latitudes coincidentes, em que prevalecem condições climáticas parecidas, geralmente têm formações vegetais com aparência semelhante e o mesmo tipo de bioma. Por exemplo, o bioma denominado Floresta Tropical predomina na faixa equatorial do planeta, tanto na América do Sul quanto na África, no sudeste da Ásia e na Oceania, embora em cada um desses locais haja espécies vegetais e animais distintas. Outro exemplo é o Cerrado brasileiro, um tipo de Savana, bioma presente em latitudes coincidentes da América do Sul, da África e da Austrália (Fig. 10).
Domínios morfoclimáticos e biomas do Brasil
Considerando características climáticas, botânicas, hidrológicas, fitogeográficas e edáficas (relativas ao solo), o geógrafo brasileiro Aziz Ab’Sáber (1924-2012) considerou a existência de seis grandes domínios morfoclimáticos em nosso país. Cada um desses domínios é caracterizado por um bioma típico, podendo apresentar outros tipos de bioma em áreas específicas. Os seis domínios morfoclimáticos propostos por Ab’Sáber são:
• Domínio Amazônico, no qual encontramos principalmente o bioma de Floresta Tropical de terra firme, bem como os biomas de Floresta de Igapó inundável e das Caatingas do Rio Negro, entre outros;
• Domínio dos Mares de Morros (ou Domínio Atlântico), cujo bioma mais típico é a Floresta Pluvial Costeira; nesse domínio também estão incluídos os biomas de Restinga e os Manguezais;
• Domínio dos Cerrados, em que predomina uma vegetação típica conhecida como Cerrado;
• Domínio das Caatingas, em que predomina um bioma de Savana Semiá- rido, incluindo também os Carnaubais;
• Domínio das Araucárias, em que predominam as Florestas de Araucária;
• Domínio das Pradarias, em que predominam os Campos Sulinos. Além desses seis domínios, há áreas de transição que abrigam biomas com características intermediárias, que não se enquadram em nenhum dos seis domínios. É o caso do Pantanal Mato-Grossense, região de transição constituída por um conjunto de diferentes biomas, entre os quais se destacam as vastas planícies inundáveis. Outros exemplos de biomas localizados em áreas de transição são as Matas de Cocais e os Manguezais. O território do Brasil, que se estende de pouco acima da linha do Equador até abaixo do Trópico de Capricórnio, favorece a existência de diversos biomas, distribuídos nos seis domínios morfoclimáticos e nas áreas de transição. Os biomas mais importantes, tanto pela área que ocupam quanto pela biodiversidade, são: Floresta Amazônica (Hileia), Floresta Pluvial Costeira (Floresta Atlântica), Floresta de Araucárias, Cerrado, Pampa, Caatinga, Floresta de Cocais (Babaçual), Manguezal, biomas que compõem o Pantanal Mato-Grossense (Figs. 11 e 12).
Figura 11 O mapa apresenta a distribuição dos domínios morfoclimáticos brasileiros e das faixas de transição.
| FIGURA 12 – LOCALIZAÇÃO DOS PRINCIPAIS DOMÍNIOS MORFOCLIMÁTICOS DO BRASIL |
A Floresta Amazônica, também conhecida por hileia amazônica, é o principal bioma do Domínio Amazônico. A Hileia se localiza na região Norte do Brasil, nos estados do Acre, Amazonas, Pará, Rondônia, Tocantins, Amapá e Roraima, além da parte norte de Mato Grosso e da parte oeste do Maranhão. Mata de igapó em Iranduba, AM, 2020.
A Floresta de Araucárias, ou Mata de Araucárias, é o principal bioma do Domínio das Araucárias, situado principalmente nos estados de Rio Grande do Sul, Santa Catarina e Paraná, em regiões com índices pluviométricos em torno de 1.400 mm anuais e temperaturas moderadas, com baixas significativas no inverno. Alguns autores referem-se a esse bioma como Floresta Subtropical Mista.
A Floresta de Araucárias, ou Mata de Araucárias, é o principal bioma do Domínio das Araucárias, situado principalmente nos estados de Rio Grande do Sul, Santa Catarina e Paraná, em regiões com índices pluviométricos em torno de 1.400 mm anuais e temperaturas moderadas, com baixas significativas no inverno. Alguns autores referem-se a esse bioma como Floresta Subtropical Mista.
O Cerrado é o principal bioma dentro do que se denomina Domínio dos Cerrados, localizado nos estados de Minas Gerais, Goiás, Tocantins, Mato Grosso, Mato Grosso do Sul, Bahia, Piauí, Maranhão, São Paulo e Paraná. Há também algumas “ilhas” dessa formação na região amazônica. Parque Nacional da Chapada dos Guimarães, MT, 2014 |
A Caatinga é o principal bioma do Domínio das Caatingas e ocupa cerca de 10% do território brasileiro, estendendo-se pelos estados de Piauí, Ceará, Rio Grande do Norte, Paraíba Pernambuco, Sergipe, Alagoas, Bahia e pelo norte de Minas Gerais. Mandacarus em Presidente Dutra, BA, 2019. |
Os Campos brasileiros, ou Pampas, compõem o Domínio das Pradarias. Os Pampas ocupam áreas de planície e localizam-se principalmente no sul do Rio Grande do Sul, caracterizando-se pela predominância de gramíneas. Campos em Quaraí, RS, 2020.
LOCALIZAÇÃO DOS BIOMAS DE TRANSIÇÃO DO BRASIL.
Em território brasileiro, o Pantanal ocupa a parte oeste dos estados de Mato Grosso e Mato Grosso do Sul, estendendo-se pelo Paraguai, pela Bolívia e pela Argentina. O Pantanal Mato-Grossense, ou Complexo do Pantanal, é um mosaico de diferentes biomas, sendo considerado uma das principais áreas de transição entre os domínios morfoclimáticos brasileiros. É uma grande planície com partes mais elevadas, denominadas informalmente de “cordilheiras”. As partes mais baixas são inundadas durante as cheias periódicas dos rios que cortam a região. Aquidauana, MS, 2018.
A Mata de Cocais, ou Floresta de Cocais ou, ainda, o Babaçual, localiza-se principalmente em certas áreas dos estados do Maranhão e Piauí. Sua espécie vegetal típica é a palmeira Orbignya martiana, o babaçu, importante economicamente para as populações locais. Das sementes da palmeira extrai-se óleo, e as folhas são utilizadas para a cobertura de casas e para a fabricação de utensílios domésticos. Nazária, PI, 2015.
O Manguezal, ou Mangue, é um bioma litorâneo presente em regiões de solo lodoso e salgado. É encontrado nas desembocaduras de rios e em áreas protegidas da ação direta do mar, como baías de águas paradas ou litorais guarnecidos por diques de areia. Durante a maré cheia, o solo do mangue fica coberto por água salobra. Os manguezais estendem-se por toda a costa brasileira, exceto nas regiões de litoral rochoso. Há mangues bem desenvolvidos no Pará, no Amazonas, no Maranhão, na Bahia, no Rio de Janeiro, em São Paulo e no Paraná. Galinhos, RN, 2020. |
|
|
|
|
|
|
|
|
|